|
این مطلب از بخش آموزش وبسایت المپیاد زیستشناسی رشد،انتخاب شده که با فرمت pdf نیز در وبسایت المپیاد رشدموجود میباشد. برای مشاهده این موضوعات در وبسایت المپیاد، به آدرس فهرست مطالب زیستشناسی مراجعه کنید. همچنین میتوانید با کلیک اینجا ، با ویژگیهای بخش آموزش این وبسایت آشنا شوید. |
تکامل رمزها
از زمانی که ساختمان رمز هر یک از اسیدهای آمینه مشخص شده است روند تکاملی این رمزها مورد سوال بوده است. چگونه 20 اسید آمینه رمزهای خود را پیدا کردهاند؟ احتمالاً زمانی که اولین جفتهای کدون و آنتیکدون (بدوی) بوجود آمدند شکلگیری رمزهای اسیدهای آمینه آغاز شد. این سیستم بتدریج روند تکاملی خویش را طی کرد و بصورت کامل درآمد.
مطالعه ردیفDNAاز ژنوم جانداران مختلف چگونگی روند تکاملی رمزها را نشان میدهد. آنچه که مسلم است جایگاه ثابت پورینها و پیریمیدینها در رمزهای پروتئین است. برای مثال رمز RNY شایعترین نوع رمز در میان اسیدهای آمینه است (که R یک پورین Y یک پیریمیدین و N میتواند هر یک از این دو باشد). و بنظر میرسد که کلیه رمزهای اولیه نیز این حالت داشتهاند. در شکل اولیه رمز RNY ممکن بود که فقط یک حالت را به خود بگیرد و یکی از سه قاب احتمالی را اختیار کند و فقط یکی از رمزهای شروع یک ژن مورد استفاده قرار گیرد.
برای مثال در میان هشت اسید آمینهای که امروزه از رمزRNY استفاده میکنند (گلیسین، ایزولوسین، ترئونین، آسپراژین، سرین، والین، آلانین و اسپارتیک اسید) احتمالاً رمزهای موجود در دوره تکاملی قبل از حیات نیز وجود داشته است که در فسیلها مشاهده شده است.
اگر امروزه ما در یک ژنوم بدنبال توالیهای تکرار شدهRNY باشیم قسمتهای کد شده و وسعت آنها را پیدا خواهیم کرد. همانطور که در شکل 1 دیده میشود،
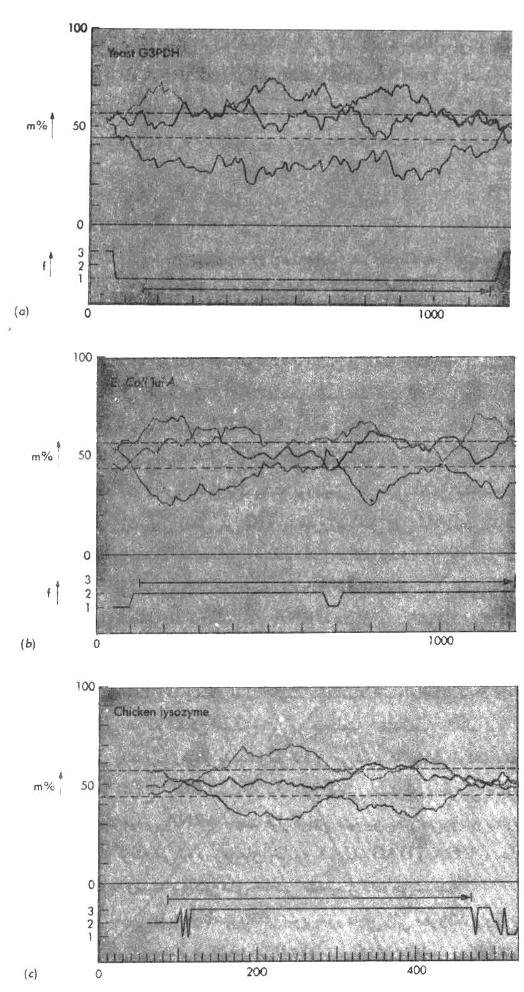 |
| شکل 1: جستجو برای یافتن پیامRNY در توالیهای DNA حاوی سه ژن. درجه موتاسیون (m%) در تکرار پذیریRNY، در مقابل عدد توالی بازی در سه قاب خواندن مختلف رسم شده است. قاب خواندنf که به بهترین نحوی با پیام RNY هماهنگ است (حداقل m% را داراست) بصورت رنگی نشان داده شده است. در هر یک از سه مورد فوق قاب و طول ناحیه کد کننده نشان داده شده است. ژن لیزوزیم جوجه در ابتدای خود بطور بدی جهش یافته است در حالیکه ژن TufA کلیباسیل ( یکی از دو ژن Ef_Tu در وسط خود دارای ناحیه کوچکی است که بنظر می رسد در آنجا تغییر قاب خواندن صورت گرفته باشد. |
ژنهایی که توانایی بیان خود را حفظ کردهاند، در حقیقت قادر بودهاند که ردیفRNYخود را دست نخورده نگهدارند (مانند آنزیم گلیسر آلدئید – 3 – فسفات دزهیدروژناز مخمر). در بعضی از ژنها ردیفهایRNY هنوز هم قابل تشخیص است اما ظاهراً در آنها موتاسیون رخ داده است (مثل آنزیم لیزوزیم مرغ) و یا اینکه در اثر موتاسیونهای از نوع حذف و یا اضافه از یک ناحیه ژن به ناحیه دیگر تغییر یافتهاند (مثل ژن TufA سلول کلیباسیل) . ژنهای نهفته باکتریوفاژ از همه جالبتر هستند، چرا که ردیف آنها در ژن اصلی انتقال یافته (ژن نهفته هنوز بصورتRNYخوانده میشود) و یا اینکه در مرحله ترجمه آنها بصورت قابهای NYRخوانده شده پروتئینهایی میسازند که درصد بسیار زیادی از اسیدهای آمینه غیر قطبی دارند. در RNY ریبوزومی 16S و یا 23S سلول کلیباسیل بدنبال قابهای تکرار شوندهRNY نظم خاصی پیدا نمیشود چونکه یا آنها اصلاً فاقد ردیفهای فوق بودهاند و یا اینکه با وجود موتاسیونهای پیاپی نظم ظاهری ردیفهایRNY بهم خورده است.
چرا قابهایRNYدر مقاطع اولیه تکاملی ابداع شد و چه اهرمهایی در انتخاب این نوع قاب نقش داشته است؟ ردیفهایRNY میتوانند در خود ساختمان مکمل تولید کنند. در ضمن نظم RNYرا در مولکولهای اولیهtRNA نیز میتوان یافت. حداقل این توجیه را میتوان کرد که مولکول های اولیه tRNA نقش دو گانه داشته و بجایmRNA نیز عمل میکردهاند بنابراین نظم ظاهری قابهایRNY در هر دو مولکول مشهور بوده است. اگر شرایط فوق مصداق داشته باشد بیان ژن قبل از پیدایش ریبوزوم توسط یک دسته از مولکولهای RNA انجام میگرفته است.