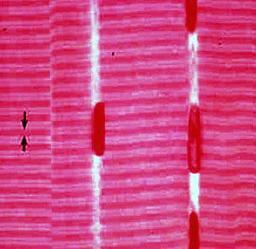
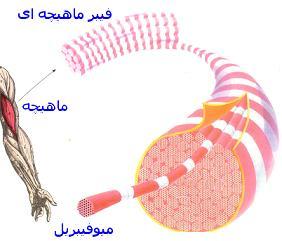
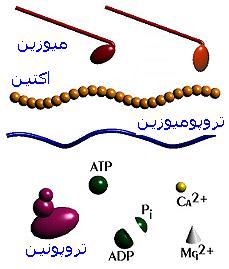
نگاه اجمالی
فیبرهای عضلات اسکلتی بوسیله فیبرهای عصبی میلیندار قطوری که از نورونهای حرکتی درشت
شاخ قدامی نخاع شروع میشوند عصبی میگردند. هر
فیبر عصبی بطور طبیعی به دفعات زیاد منشعب شده و از 3 تا چندصد فیبر عضله اسکلتی را تحریک می کنند. انتهای رشتههای عصبی یک محل تماس با
فیبر عضلانی تقریبا در وسط آن به نام محل تماس عصبی عضلانی برقرار میکند بطوری که پتانسیل عمل در هر دو جهت به سوی انتهای فیبر عضلانی سیر میکند. به استثنای دو درصد فیبرهای عضله ، فقط یک محل تماس عصبی عضلانی برای هر فیبر عضلانی وجود دارد.
شریح فیزیولوژیک محل تماس عصبی عضلانی
فیبر عصبی در انتهای خود منشعب شده و مجموعهای از ترمینالهای منشعب عصبی تشکیل میدهند که در یک فرو رفتگی از سطح فیبر عضلانی قرار میگیرد اما بطور کامل در خارج
غشای پلاسمایی فیبر عضلانی قرار دارد. تمام این تشکیلات صفحه حرکتی انتهایی نامیده میشود و بوسیله یک یا چند
سلول شوان پوشیده میشود که آن را از
مایعات اطراف مجزا میسازد.
فرورفتگی غشا فیبر عضلانی موسوم به
ناودان سیناسپی و فضای بین ترمینال عصبی و غشای فیبر عضلانی موسوم به شکاف سیناسپی (Synaptic Cleft) است. پهنای شکاف سیناسپی 20 تا 30 نانومتر بوده و توسط لایه نازکی از فیبرهای مشبک اسفنجی موسوم به تیغه پایه اشغال میشود که مایع خارج سلولی از میان آن انتشار مییابد. در عمق ناودان سیناسپی چین خوردگیهای کوچک متعددی در غشای فیبر عضلانی موسوم به شکافهای زیر عصبی وجود دارند که وسعت سطحی را میانجی سیناسپی در آن عمل کند به مقدار زیادی افزایش میدهد.
در انتهای آکسون
میتوکندریهای متعددی وجود دارند که بطور عمده تامین کننده انرژی جهت سنتز میانجی تویکلی استیل کولین هستند. استیل کولین به نوبه خود فیبر عضلانی را تحریک میکند. استیل کولین در
سیتوپلاسم انتهای
فیبر عصبی ساخته میشود. اما به سرعت جذب تعدا زیادی
وزیکولهای سیناسپی کوچک میگردد که تقریبا 300000 عدد از آنها بطور طبیعی در ترمینالهای یک صفحه حرکتی انتهایی واحد وجود دارند چسبیده به ماتریس تیغه پایه مقادیر زیادی از آنزیم کولین استراز وجود دارد که میتواند استیل کولین را منهدم کند.
ترشح استیل کولین توسط ترمینالهای عصبی
هنگامی که یک ایمپالس عصبی به محل تماس عصبی عضلانی میرسد حدود 125 ویزیکول استیل کولین بوسیله ترمینالها به داخل شکاف سیناسپی آزاد میشوند. در سطح داخلی غشای عصبی میلههای متراکم مستقیم وجود دارد. در طرفین هر میله متراکم ذرات پروتئینی وجود دارند که به داخل غشا نفوذ میکنند و معتقدند که کاناهای کلسیمی دریچهدار ولتاژی هستند. هنگامی که پتانسیل عمل در سراسر ترمینال پخش میشود این کانالها باز شده و اجازه میدهند تا یون
کلسیم به مقدار زیاد داخل ترمینال انتشار یابد.
معتقدند که یونهای کلسیم به نوبه خود یک اثر جاذبهای به روی فیزوکولهای استیل کولین اعمال کرده و آنها را به سوی
غشای عصبی مجاور میلههای متراکم میکشند. پارهای از ویزیکولها با غشای عصبی جوش میخورند و استیل کولین خود را توسط روند اگزوسیتوز به داخل شکاف سیناسپی آزاد میکنند. اگرچه بعضی از جزئیاتی که در بالا ذکر شد هنوز جنبه فرضی دارند. اما معلوم شده که
اتیمولوس موثر برای ایجاد استیل کولین آزاد شد. از وزیکولها ، ورود یونهای کلسیم است. علاوه بر آن ، ویزیکولها در محلی از غشا که در مجاورت میلههای متراکم قرار دارد، تخلیه میشوند.
اثر استیل کولین بر غشا سیناسپی
رسپیتورهای استیل کولینی در غشا عضله در واقع کانالهای یونی دریچهدار وابسته به
استیل کولین هستند که تقریبا بطور کامل در نزدیکی دهانههای شکافهای زیر عصبی بلافاصله در زیر میلههای متراکم قرار گرفتهاند که در آنجا ویزیکولهای استیل کولین به داخل شکاف سیناسپی تخلیه میشوند. هر رسپیتور یک مجموعه پروتئینی درشت با وزن مولکولی کل 275000 بوده و از پنج زیر واحد پروتئینی تشکیل شده که در سراسر عرض غشا نفوذ میکنند و بطور دایرهوار در کنار یکدیگر قرار میگیرند و یک کانال لولهای تشکیل میدهند. کانال به حالت بسته باقی میماند تا اینکه دو استیل کولین به دو زیر
واحد پروتئینی بچسبد.
این امر موجب بروز یک تغییر شکل فضایی میشود که کانال را باز میکند. کانال استیل کولینی در هنگام باز بودن قطری حدود 65% نانومتر دارد که آنقدر بزرگ است که به تمام یونهای مثبی مهم نظیر یون
سدیم ، یون
پتاسیم و یون
کلسیم اجازه میدهد تا به آسانی از منفذ کانال عبور کنند. از طرف دیگر ، یونهای منفی از قبیل یونهای
کلر به علت بارهای منفی قوی در دهانه کانال از این کانال عبور نمیکنند. در عمل یونهای سدیم بسیار بیشتری از هرگونه یون دیگری به دو دلیل زیر از کانالهای استیل کولینی جریان مییابند:
- فقط دو یون مثبت با غلظت به اندازه کافی زیاد وجود دارند که از این نظر میتوانند اهمیت داشته باشند که عبارتند از یونهای سدیم در مایع خارج سلولی و یونهای پتاسیم در مایع داخل سلولی
- پتانسیل بسیار منفی در داخل غشای عضله یعنی 80- تا 90- میلی ولت یونهای با بار مثبت سدیم را به داخل فیبر میکشاند در حالیکه همزمان با آن در صورتی که یونهای پتاسیم کوشش کنند که به طرف خارج بروند از خروج یونهای پتاسیم جلوگیری میکند.
بنابراین اثر عمده باز شدن کانمالهای دریچهدار استیل کولینی اجازه دادن به تعداد زیادی از یونهای سدیم برای ریختن به داخل فیبر است که همراه با خود تعدا زیادی از بارهای مثبت را حمل میکنند. این امر یک تغییر پتانسیل موضعی در غشای فیبر عضلانی موسوم به پتانسیل صفحه انتهایی تولید میکند. این پتانسیل صفحه انتهایی به نوبه خود موجب بروز یک پتانسیل عمل در غشای عضله میشود و به این ترتیب باعث انقباض عضله میگردد.
انهدام استیل کولین آزاد شده توسط استیل کولین استواز
استیل کولین پس از آزاد شدن به داخل فضای سیناسپی تا زمانی که در این فضا باقی بماند به فعال کردن رسپتورهای استیل کولینی ادامه خواهد داد. اما باید دانست که استیل کولین به سرعت از دو راه حذف میگردد.
- قسمت اعظم استیل کولین توسط آنزیم استیل کولین استواز منهدم میشود که بطور عمده به تیغه پایه میچسبد که یک لایه اسفنجی از بافت همبندی ظریف است که فضای سیناسپی بین ترمینال پیش سیناسپی و غشای عضلانی پس سیناسپی را پر میکند.
- مقدار کمی از آن به خارج از شکاف سیناسپی انتشار مییابد و در این حال دیگر در دسترس قرار ندارد تا بر روی غشای فیبر عضلانی عمل کند. مرحله زمانی بسیار کوتاهی که استیل کولین در شکاف سیناسپی باقی میماند (حداکثر چند میلی ثانیه) تقریبا همیشه برای تحریک فیبر عضلانی کافی است، سپس حذف سریع استیل کولین از تحریک مجدد عضله بعد از آنکه فیبر از پتانسیل عمل اول خارج شد جلوگیری می کند.
مباحث مرتبط با عنوان