فقدان یک اپراتور منجر به سنتز دائمی یک پروتئین میشود
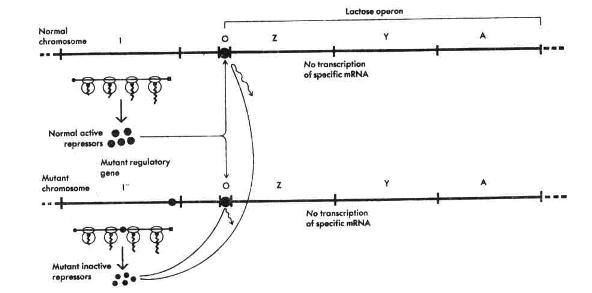
اپراتورها که به آنها مولکولهای سدکننده وصل میشوند نیز مانند سدکنندهها بطور معکوس عمل میکنند و بدین معنی که در صورت عدم عمل اپراتور، سدکننده مربوط به آن نمیتواند سنتزmRNA مخصوص آنرا مهار کند، در نتیجه محصولات پروتئینی مزبور بطور دائم سنتز خواهند شد.
وجود اپراتورها اولینبار با بررسیهای ژنتیکی، به اثبات رسید. یک اپراتور ممکن است غیرفعال شود، در نتیجه سدکننده نمیتواند به آن وصل شود. در چنین صورتی آنزیمها بطور دائم سنتز میشوند. این جهشیافتهها بنام

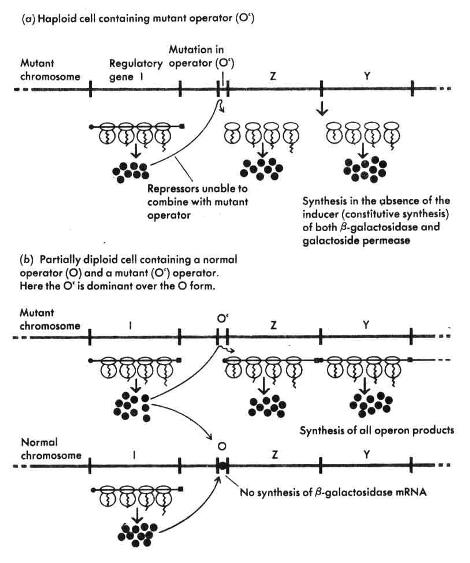
خوانده شده، براحتی از جهش یافتههایی که اشکالی در تولید سدکننده دارند، تمیز داده میشوند. این تمایز با استفاده از سلولهایی که دیپلوئید جزیی هستند یعنی تنها قسمتی از کروموزوم آنها دو نسخهای است انجام میشود. در سلولی که یک ژن سدکننده لاکتوز فعال و یک ژن غیرفعال وجود داشته باشد،اپرون لاکتوز قابل سد شدن است چرا که مولکولهای سدکننده سالم مربوط به ژن سالم میتوانند با هر دو اپراتور واکنش دهند (شکل1 ).
| شکل1 : از سلولهایی که دیپلوئید جزیی هستند برای نشان دادن سدکننده سالم حتی در صورت وجود یک سدکننده غیرفعال استفاده میشود. در چنین سلولهایی در صورت وجود بتاگالاکتوزیدها، بتاگالاکتوزیداز زیادی تولید نمیشود. |
از طرف دیگر در سلولهایی که تنها یک اپراتور وجود داشته باشد، صرفنظر از شرایط ژن سدکننده، آنزیمهای اپرون لاکتوز بطور دائم سنتز میشوند (شکل2 ).
|
شکل2 : چگونگی کنترل سنتز یک mRNA اختصاصی در اپراتورهای طبیعی و جهش یافته |
کنترل مثبت اپرون لاکتوز
با وجودیکه ابتدا تصور میشد که اپرون لاکتوز تنها تحت کنترل منفی سدکننده لاکتوز است ولی امروزه میدانیم که القاکننده مثبتی وجود دارد که عمل سدکننده را خنثی میکند. کنترل مثبت فوق با واسطه پروتئینی که برای بیان طبیعی اپرون لازم است، صورت میگیرد. وجود این پروتئین اولین بار در هنگام آزمایشاتی که نحوه بلوکه کردن بعضی از اپرونها بوسیله گلوکز را نشان میداد، کشف شد. اپرونهای فوق که بنام اپرونهای حساس به گلوکز خوانده میشوند، هر یک از کاتابولیسم یک قند بخصوص (مثلاً گالاکتوز، لاکتوز، آرابینوز و مالتوز) را کنترل میکنند. برای مثال هنگامی که کلیباسیل در حضور گلوکز و لاکتوز رشد یابد، تنها گلوکز مصرف میشود و محصولات اپرون لاکتوز (سه پروتئین اختصاصی) ساخته نمیشوند. همچنین در شرایطی که گلوکز و گالاکتوز وجود داشته باشند، اپرون غیرفعال است. شاید دلیل ترجیح گلوکز در چنین شرایطی یک علت تکاملی داشته است، بدین معنی که باکتریها در طول تکامل همواره در محیط غنی از گلوکز بوده، با قندهای دیگر کمتر در تماس بودهاند.
مقدارAMPحلقوی تحت تاثیر میزان کاتابولیسم گلوکز قرار میگیرد
رونویسی اپرون لاکتوز بطور مستقیم بوسیله گلوکز کنترل نمیشود، بلکه در اثر کاهش درون سلولی مقدار AMPحلقوی (cAMP) که یکی از کاتابولیتهای گلوکز است تنظیم میشود. رونویسی از کلیه اپرونهایی که در اثر کاتابولیسم گلوکز مهار میشوند، مستلزم وجودcAMP است. هنوز معلوم نیست که کاتابولیتهای گلوکز چگونه مقدارcAMP را کنترل میکنند. این ترکیب بطور مستقیم بوسیله آنزیمی بنام آدنیلات سیکلاز از ATP بوجود میآید. آنزیم فوق ممکن است مستقیماً بوسیله یک کاتابولیت اختصاصی گلوکز مهار شود (شکل3 ).
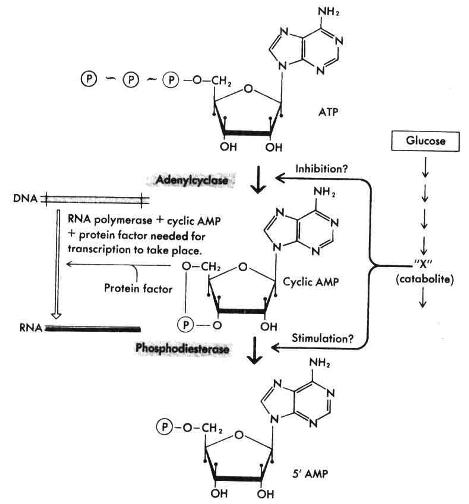 شکل3 : کنترل رونویسی اپرونهای حساس به CAP از طریق cAMP. |
از طرف دیگر آنزیم دیگری بنام فسفودی استراز وجود دارد که بطور اختصاصیcAMP را به AMP تبدیل میکند. کاهشcAMP درون سلولی ممکن است با افزایش سرعت تخریب cAMP صورت گیرد.
پروتئین فعالکننده ژن کاتابولیت (CAP) در اثر اتصال به cAMP فعال میشود
cAMP بطور مستقیم باعث افزایش سنتزmRNA اپرون لاکتوز نمیشود بلکه ابتدا به پروتئینی بنام فعالکننده ژن کاتابولیت(CAP) متصل میشود.CAP مولکول دیمری به وزن مولکولی 45000 دالتون است. CAPبه تنهائی هیچ تاثیری بر رونویسی ندارد، مگر آنکهcAMP قبلاً به آن متصل شده باشد. مجموعه فوق با اتصال به جایگاههای بسیار اختصاصی درDNA، باعث افزایش رونویسی اپرونهای مجاور میشود.CAP، یک عنصر کنترلی مثبت برای کلیه اپرونهای حساس به گلوکز است. بنابراین در سلولهایی که ژنCAP دچار جهش شده باشد، قندهای زیادی نمیتوانند مصرف شوند.
پیوند های خارجی
http://Olympiad.roshd.ir/biology/content/pdf/0144.pdf