مقدمه
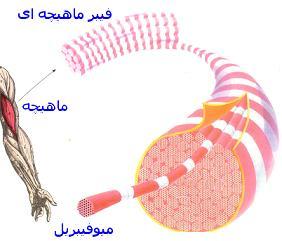
این نوع عضله را به خاطر داشتن نوارهای تیره و روشن در زیر میکروسکوپ عضله مخطط ، به علت چسبیده بودن آنها به استخوانها ، عضله اسکلتی و به جهت عملکرد ارادی آنها ، عضلات ارادی مینامند که وظیفه اصلی این نوع عضله شرکت در حرکات بدن میباشد. سلولهای عضله مخطط چون از بهم پیوستن تعداد زیادی سلول سازنده عضلانی که
میوبلاست نام دارد بوجود میآیند، بسیار بلند هستند و رشتههای عضلانی که نیز نامیده میشوند. گروهها سلولهای عضلانی را که از تعدادی سلول موازی هم در هر گروه تشکیل میگردد دسته و مجموعه دستهها را باهم عضله مینامند.
ساختمان میکروسکوپی عضله مخطط
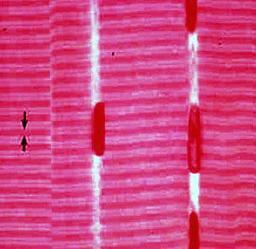
با میکروسکوپ نوری در مقطع طولی عضله مخطط دو نوع نوار عرضی دیده میشود که این نوارها با توجه به خصوصیات فیزیکی آنها به دو دسته تقسیم میشوند. نوارهایی که در برابر نور پلاریزه دارای خاصیت انکسار مضاعف هستند که به
نوارهای (Aisotropic) یا A و نوارهایی که در مقابل نور پلاریزه فاقد این خاصیت هستند. (Isotropic) نوارهای آنیزوتروپیک را نوارهای تیره و نوارهای ایزوتروپیک را نوارهای روشن مینامند.
در مرکز نوار روشن نوار تیره بسیار ظریفی دیده میشود که
نوار Z نام دارد. حد فاصل بین دو نوار Z را سارکومر مینامند که شامل یک نوار A و نیمی از نوارهای روشن در طرفین آن میباشد. سارکومر واحد ساختمانی و فیزیولوژیکی سلول عضلانی محسوب میشود. با میکروسکوپ الکترونی در قسمت میانی نوار تیره ناحیه نسبتا روشنی دیده میشود که به نوار H موسوم است و در مرکز
نوار H خط باریک و تیرهای مشاهده میشود که به
خط M موسوم است.
ساختمان مولکولی میوفیبریلها
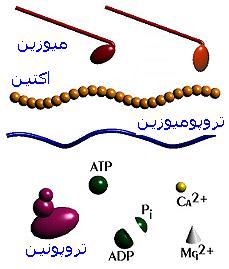
میوفیبریلها یا میوفیلامنتها شامل فیلامنتهای نازک و ضخیم است که بر روی هم 55 درصد کل پروتئین عضله را تشکیل میدهند.
فیلامنتهای نازک
فیلامنتهای نازک در سلولهای عضله مخطط حاوی سه نوع پروتئین به اسامی
اکتین ،
تروپونین و
تروپرومیوزین میباشد.
فیلامنتهای ضخیم
فیلامنتهای ضخیم از
میوزین تشکیل شدهاند. میوزین مولکول بزرگی است به طول 200 نانومتر که از دو قسمت سر و دنباله تشکیل شده است.
مکانیسم انقباض
انقباض عضله در اثر اتصال سر میوزین به اکتین ، فعال شدن ATPase و تجزیه
ATP ، خم شدن سر میوزین و کشیده شدن فیلامنتهای اکتین به حد فاصل فیلامنتهای میوزین و کوتاه شدن سارکومر انجام میشود. این فرضیه که اولین بار توسط
هاکسلی مطرح گردید به فرضیه لغزیدن فیلامنت نیز موسوم است. در عضله در حال استراحت محل قابل اتصال اکتین با میوزین توسط مجموعه تروپونین _تروپومیوزین پوشیده شده و مانع اتصال میوزین به اکتین میشود.
در موقع انقباض ، تحریکات عصبی موجب خروج کلسیم از ذخایر درون سلولی و افزایش غلظت داخل سیتوپلاسمی کلسیم میشود. به دنبال این فرایند کلسیم به تروپونین چسبیده و با ایجاد تغییر شکل فضایی در مولکول تروپونین سبب جابجایی تروپومیوزین و آشکار شدن ناحیه قابل اتصال اکتین با میوزین میشود.
رگها و اعصاب عضلات مخطط
عصبی که به عضلات مخطط وارد میشود حاوی رشتههای حسی و حرکتی است. رشتههای عصبی در این عضله از نوع میلیندار و بدون میلین است.
ترمیم عضله اسکلتی
عواملی مانند قطع عصب ، نرسیدن مواد غذایی و التهاب موضعی میتوانند باعث لاغری و یا مرگ سلول عضلانی شوند. سلولهای عضله مخطط غیر قابل تقسیم هستند و قادر به ترمیم سلولهای از بین رفته نمیباشند. در نتیجه عمده سلولهای از بین رفته ، بوسیله
بافت همبند و
چربی جایگزین میگردد. با وجود این ترمیم محدود بافت عضلانی در اثر تکثیر و تمایز سلولهایی به نام سلولهای قمری امکان پذیر است.
ترشح استیل کولین توسط ترمینالهای عصبی
هنگامی که یک ایمپالس عصبی به محل تماس عصبی عضلانی میرسد حدود 125 وزیکول استیل کولین بوسیله ترمینالها به داخل شکاف سیناسپی آزاد میشوند. در سطح داخلی غشای عصبی میلههای متراکم مستقیم وجود دارد. در طرفین هر میله متراکم ذرات پروتئینی وجود دارند که به داخل غشا نفوذ میکنند و معتقدند که کاناهای کلسیمی دریچهدار ولتاژی هستند. هنگامی که پتانسیل عمل در سراسر ترمینال پخش میشود این کانالها باز شده و اجازه میدهند تا یون
کلسیم به مقدار زیاد داخل ترمینال انتشار یابد.
معتقدند که یونهای کلسیم به نوبه خود یک اثر جاذبهای به روی وزیکولهای استیل کولین اعمال کرده و آنها را به سوی
غشای عصبی مجاور میلههای متراکم میکشند. پارهای از وزیکولها با غشای عصبی جوش میخورند و استیل کولین خود را توسط روند اگزوسیتوز به داخل شکاف سیناسپی آزاد میکنند. اگرچه بعضی از جزئیاتی که در بالا ذکر شد هنوز جنبه فرضی دارند. اما معلوم شده که اتیمولوس موثر برای ایجاد استیل کولین آزاد شد. از وزیکولها ، ورود یونهای کلسیم است. علاوه بر آن ، وزیکولها در محلی از غشا که در مجاورت میلههای متراکم قرار دارد، تخلیه میشوند.
مباحث مرتبط با عنوان